太阳光不仅是植物生长的能量来源,还是调节植物生长发育过程的关键信号。光敏色素是植物的红光/远红光感受器,作为植物的“眼睛”扮演重要角色。当下全球气候变化愈加频繁、耕地资源逐步缩紧,维护全球粮食安全是一严重挑战。过去近一个世纪,农作物产量的不断提高得益于越来越耐密植的作物新品种。将来作物产量进一步提高也将继续依赖于培育更耐密集种植的新品种。由于密植遮荫环境中植物的感光机理和耐密高产性状均和植物光敏色素蛋白息息相关,阐明植物光敏色素响应以及传递光信号的机制将有助于改善作物密植性能及对复杂环境的适应性,从而为粮食安全的保障做出贡献。
高等植物主要编码以光敏色素A(phyA)和光敏色素B(phyB)为代表的两类光敏色素,其中phyB是介导可逆红光响应的主要红光受体。光敏色素通过发色团PΦB在红光吸收态(Pr,基态)以及远红光吸收态(Pfr,激活态)之间进行可逆转变。拟南芥phyB被光激活后,可以直接与一类光敏色素互作因子(phytochrome-interacting factor,PIF)互作,传递光信号并调控下游基因表达,促进光形态建成。因此phyB和PIFs构成了植物响应周围光环境的关键信号模块。拟南芥中8个PIF成员(PIF1-8)均包含两个重要结构域:N端为结合phyB-Pfr的激活态结合域(Active-PHYB Binding motif,APB),其C端为结合DNA的bHLH二聚化结构域。在黑暗条件下,phyB-Pr定位于细胞质中,PIF1/3/4/5在核中作用驱动幼苗暗形态建成的发育过程(下胚轴伸长,子叶闭合)。一旦幼苗感知红光,phyB-Pfr入核进而负调控上述PIFs,抑制下胚轴伸长和促进子叶展开,并维持植物光生长形态。尽管2022年美国Vierstra团队在Nature发文揭示了phyB-Pr的结构,但phyB-Pfr及其识别PIF的结构生物学基础及其调控机理仍不清楚。
2024年9月24日,北京大学现代农学院王继纵课题组与邓兴旺课题组合作在Cell发表了题为Light-induced remodeling of phytochrome B enables signal transduction by phytochrome-interacting factor的研究论文,揭示了长期以来期待的phyB光信号转导的最初反应机制。

该研究解析了模式植物拟南芥光激活态phyB-Pfr以及不依赖于光的组成型激活突变体phyBY276H分别结合下游信号分子PIF6的复合物高分辨率冷冻电镜结构(分辨率依次为3.1 Å,3.2 Å)。基于分子结构分析、结合突变体蛋白光谱性质测定、体外以及半体外生化实验表征、以及转基因植物的表型分析,揭示了光激活phyB由Pr转变为Pfr的详细分子机制,并提出了phyB和PIF之间“诱导-契合”的相互作用模型,填补了植物光敏色素信号传导机制研究的一个关键空白。
由于全长PIFs蛋白存在大量无序结构,体外重组表达以及组装phyB-PIF全长蛋白复合物存在相当大的困难。考虑到系列文献报道中PIFs的APB motif(N端100个氨基酸)足以介导其与phyB在体内及体外互作,研究者选取了结合能力最强的PIF6-APB(PIF6-100,仅N端100个氨基酸)进行phyB-PIF的复合物组装并制备冷冻样品。最终成功解析得到了phyB-Pfr-PIF6以及缺失HKRD结构域(仅含N端908个氨基酸)的phyBY276H-908-PIF6复合物的高分辨率冷冻电镜结构。结构分析表明phyB-Pfr在光激活之后发生大规模结构重排,其光感应模块(photosensory module, PSM)从Pr状态下“头对尾”转变为Pfr状态下“头对头”二聚体,且PIF6-APB单体结合在phyB-Pfr二聚体界面的一侧,组成phyB-PIF6三聚体。此外,phyBY276H-908-PIF6复合物结构与phyB-Pfr-PIF6几乎完全一致(图1)。
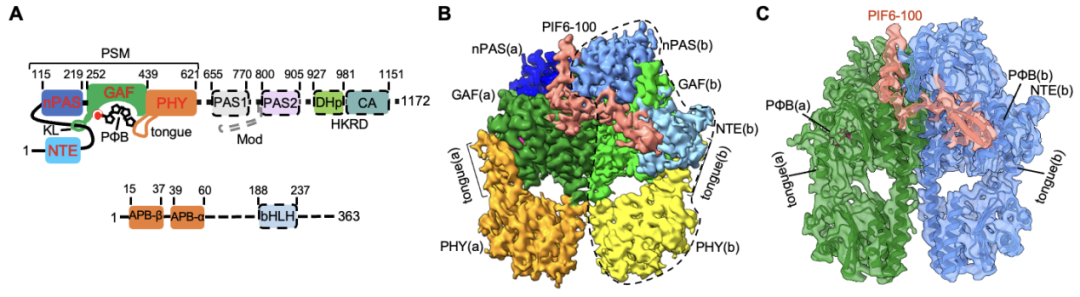
图1. phyB-Pfr-PIF6(B)和phyBY276H-PIF6(C)均为不对称三聚体
为了阐明光激活驱动phyB的结构重排,作者细致比对了之前文献报道的phyB-Pr及本研究获得的phyB-Pfr结构,发现发色团PΦB分子在吸收红光后,其D环翻转了180˚,并且和口袋中的系列氨基酸重新建立互作网络(图2A,左),最终导致PHY结构域里与其互作的舌状突出结构发生由β片层到α螺旋的构象转变(图2A,右)。结构分析表明,丝氨酸S584对于稳定α螺旋形式的舌状结构非常重要(图2A,右),进一步的体外生化以及转基因植物表型分析证明了S584对于维系phyB-Pfr激活状态和phyB信号通路至关重要(图2B)。

图2. 光诱导PΦB分子(A,左)和PHY结构域舌状突出结构(A,右)的构象转变及功能验证(B)
在Pr状态下,phyB的C末端结构域PAS2以及HKRD与N末端结构域GAF和PHY互作以维系“头对尾”二聚体构象。而在phyB-Pfr中,其PHY结构域的舌状突出结构转变为α螺旋后,会直接和Pr状态里的PAS2结构域发生空间冲突(图3A,左;图3B,左),从而破坏Pr状态下PAS2和PHY结构域之间的广泛互作,这会进一步破坏HKRD和PHY结构域(图3A,左;图3B,中),以及HKRD和GAF结构域(图3A,右;图3B,右)之间的分子内互作,这一系列的构象变化完全打破Pr状态下的“头对尾”结构,最终激活phyB。此外,缺失HKRD结构域的phyB截短体蛋白(phyB-908,仅含N端908个氨基酸)在光激活之后倾向于单体形式存在(图3C),进一步佐证了phyB-Pr在光激活后,原来由N端和C端模块互作维系的“头对尾”二聚体被打破,进而形成phyB-Pfr。
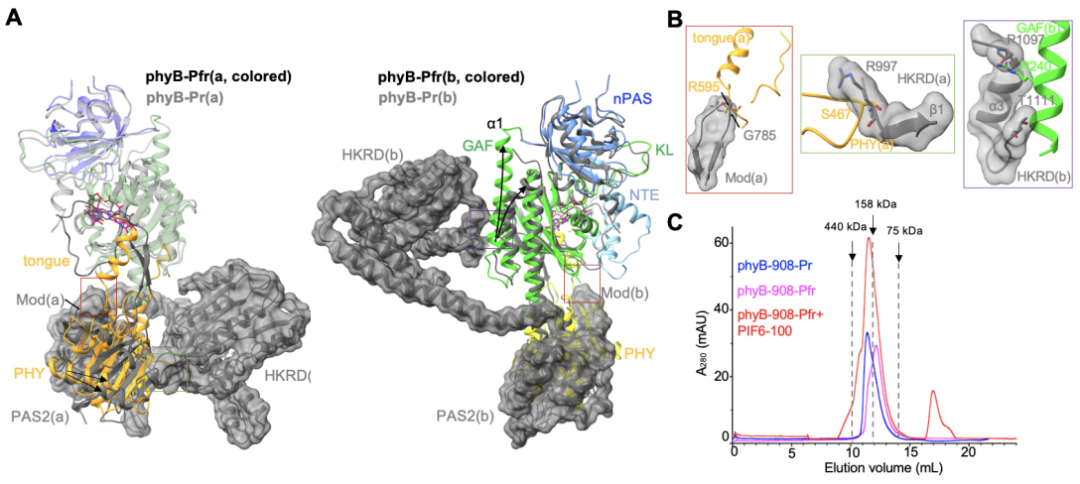
图3. PHY 结构域舌状突出结构的构象转变驱动phyB-Pr重构形成phyB-Pfr(A,B)及生化验证(C)
光激活后的phyB-Pfr分别利用其N末端的NTE结构域以及由整体N末端结构域形成的“头对头”二聚体,来识别和结合PIF6-APB的N末端和C末端(图4A)。PIF6-APB的N末端通过形成一个β发夹结构(PIF6-APB-β)与phyB-Pfr的NTE结构域发生广泛互作(图4B),而phyB-Pfr二聚体结构中另一分子phyB并不具备稳定结构的NTE,表明NTE的稳定结构是由PIF6-APB的结合所诱导形成的。此外,PIF6-APB的C末端通过形成一个α螺旋结构(PIF6-APB-α)同时与两个phyB-Pfr的N末端结构域产生广泛互作(图4C),印证了PIF6-100促进的phyB-908-Pfr的二聚化(图3C)。作者进一步通过系列生化、光谱实验表征、结合转基因植物的表型明确了NTE结构域以及N末端结构域介导的phyB-Pfr二聚化对于phyB-PIF互作的重要性。
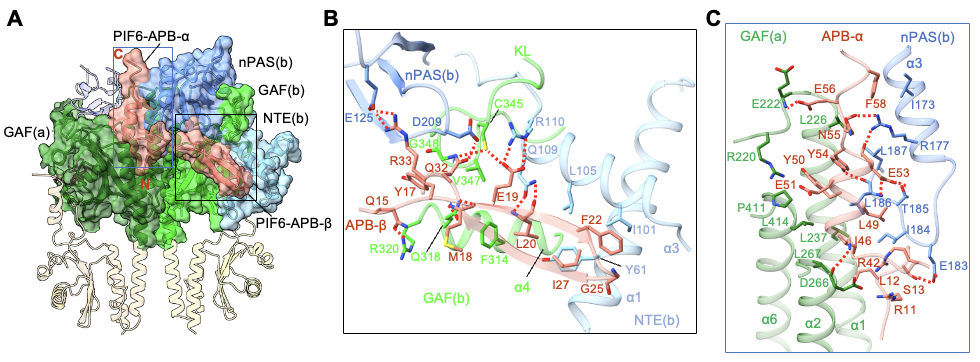
图4. phyB-Pfr的N末端结构域对PIF-APB的特异性识别(A)以及NTE结构域特异性识别PIF-APB的N末端β发夹结构(B),phyB-Pfr二聚体特异性识别PIF-APB的C末端α螺旋结构(C)
两个phyB-Pfr的结构比对表明phyB-PIF6复合物结构中的phyB-Pfr形成了不对称的二聚体(图5A),这些不对称性导致phyB-Pfr二聚体的另一侧界面没有足够空间与PIF6-APB-β以及PIF6-APB-α形成合适的互作(图5B)。作者进一步通过pull-down以及Co-IP assay评估了溶液状态下phyB和PIF-APB以及PIF-FL的互作模式,生化数据表明PIF3和PIF6的APB motif以及PIF1和PIF3的全长蛋白均可以和phyB-Pfr二聚体形成摩尔比为1:2(2:4)的复合物。
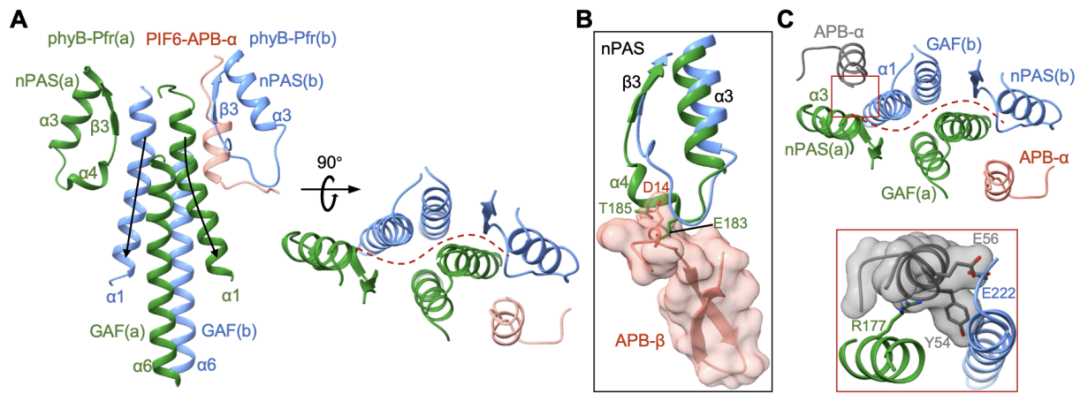
图5. 不对称的phyB-Pfr二聚体只能结合一个PIF6-APB单体
总的来说,这项研究揭示了“头对头”激活态phyB二聚体识别结合PIF6-APB的复合物结构,解决了光敏色素研究中光如何诱导phyB变构激活并转导PIF信号的关键问题(图6),为作物感光性状的改造以及phyB相关光控基因表达的“光遗传学”工具开发提供了分子水平的精细图纸。
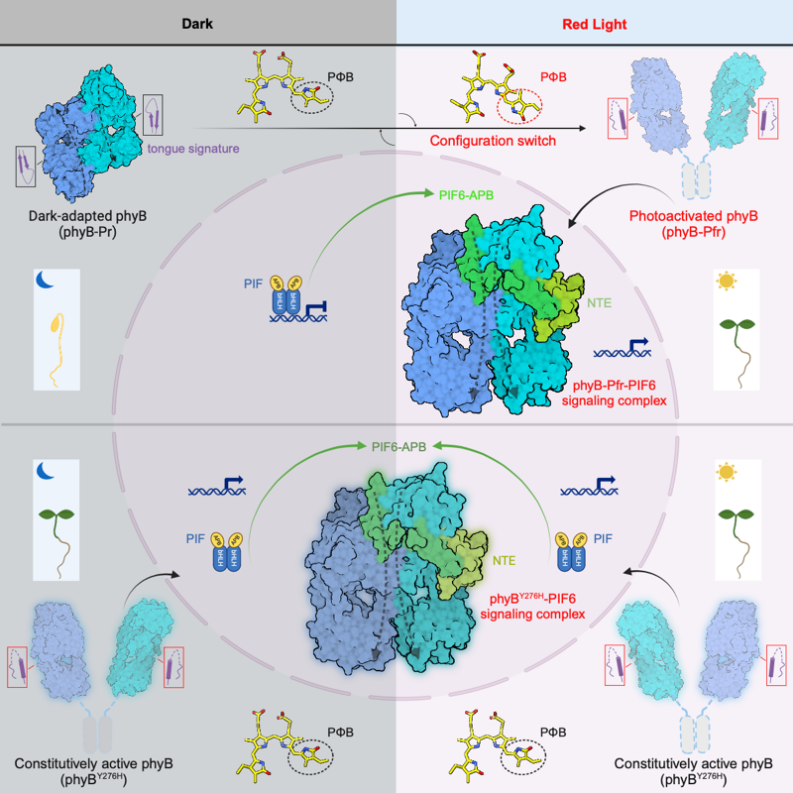
图6. 红光激活phyB并转导PIF信号的结构机制模型
北京大学现代农学院、北京大学现代农业研究院王继纵研究员和邓兴旺教授为该论文的通讯作者。北京大学前沿交叉学科研究院博士研究生王征东和宋艳萍、北京大学现代农业研究院科研助理王文凤和赵迪迪、以及北京大学现代农业研究院林晓莉博士为论文共同第一作者。北京大学现代农业研究院赵珺博士、迟程博士和高级工程师徐斌、中国科学院遗传与发育生物学研究所博士研究生沈萌对本研究也做出了重要贡献。冷冻样品制备、样品筛选和数据收集在研究院生物微观结构研究平台完成。该研究得到了国家自然科学基金、国家重点研发项目、山东省重点研发项目、山东省科技创新基金、中科协青年人才托举工程、山东省泰山青年学者项目、北京大学现代农业研究院、潍坊现代农业山东省实验室、小麦育种全国重点实验室、北京大学蛋白质与植物基因研究国家重点实验室相关经费的资助。
论文链接:https://www.cell.com/cell/fulltext/S0092-8674(24)01023-7
专家观点
万建民 院士(中国农业科学院作物科学研究)
光照是植物生长的能量来源,对农作物产量起着决定性作用。在农业生产中,提高作物种植密度是增产的重要途径。然而,随着密度增加,作物植株之间相互遮阴会加剧,造成植株茎秆增高、变弱,易发生倒伏、小穗和抗逆性减弱等突出问题。因此,阐明植物感知光照的分子机理对提高作物光适应能力和耐密性等具有十分重要的意义。光敏色素是植物的红光、远红光受体,是影响作物响应光照变化的关键功能蛋白。自1960年首次以“光敏色素”命名这类光受体以来,研究人员一直好奇光敏色素感知红光/远红光信号的机制。早在上世纪80年代末,研究人员在植物光敏色素phyA的电镜照片中就看到了“Y”字形的蛋白颗粒。但是由于光敏色素属于多结构域光受体蛋白,植物内源含量较低,且重组表达困难,研究人员主要通过对相对容易获得的微生物来源光敏色素进行结构表征,植物光敏色素的结构研究相对停滞。直到2014年,PNAS杂志发表了基态phyB光感应模块截短体的晶体结构,2022年以来,来自于美国以及中国的多家单位相继报道了全长phyB及phyA基态的结构,从而逐渐建立了光敏色素基态结构特性的认识。但植物光敏色素激活态结构及光信号原初转导的机制仍不清楚,限制了基于激活态phyB结构的光信号转导机制研究。
这项研究首次报道了激活态植物光敏色素phyB结合标志性下游信号分子PIF的复合物结构,揭示了广泛研究的不依赖与光照的组成型激活突变体phyBY276H结合PIF的复合物结构,并结合多种研究方法,阐明了光照引起的激活态phyB结构重排以及光敏色素蛋白和发色团分子之间相互协调的动态变化机制。这项研究充分回答了长期以来研究人员关注的phyB感光激活机理,填补了激活态植物光敏色素结构研究的空白,是生物学基础研究的重大突破。同时,光敏色素激活机制的解析为精准调控植物光响应建立了基础,对定向改良玉米、小麦、水稻等重要作物的感光性状、培育耐密高产品种意义重大。
王海洋 教授(华南农业大学)
培育耐密植作物是提高作物单产的有效技术措施。株型是决定作物耐密性的关键因素之一。然而,密植容易诱发植物产生避荫反应,导致植株徒长、茎秆变细、根系发育减弱、增加倒伏风险等不利后果。作为植物响应红光/远红光信号的主要红光受体,光敏色素phyB对于植物应对外界光环境变化的适应性生长(株高、叶夹角、开花时间等)至关重要,从而影响作物的株型、光合效率和产量。phyB被红光激活后,通过与PIF蛋白直接互作传递光信号,进而启动和维持光形态建成的发育过程。然而自1998年发现PIF是激活态光敏色素结合蛋白以来,虽然鉴定了部分影响phyB-PIF互作的关键位点,但激活态phyB与PIF蛋白互作的结构基础和调控机理目前仍然未知。这项研究通过解析了模式植物拟南芥光激活态phyB-Pfr以及不依赖于光的组成型激活突变体phyBY276H分别结合下游信号分子PIF6的复合物高分辨率冷冻电镜结构,揭示了phyB基态(Pr)在光激活后,其光感应模块(PSM)从Pr状态下“头对尾”转变为Pfr状态下“头对头”二聚体。由于phyB-Pfr二聚体的不对称性,导致其仅一侧能够结合PIF并形成phyB-Pfr-PIF的三聚体。作者进一步结合突变体蛋白光谱性质测定、体外以及半体外生化实验表征、以及转基因植物的表型分析,验证了丝氨酸S584、NTE结构域以及N末端结构域对于介导phyB-Pfr二聚化,phyB-PIF互作和phyB信号通路的重要性。该研究首次揭示了phyB光激活及最初光信号传递(与PIF互作)的结构生物学基础,是近年来植物光敏色素信号传导研究领域取得的又一个重大突破。研究成果可为培育耐荫、耐密、抗倒伏、高光效作物新品种提供重要理论指导和基因资源。